Биогеография океанов и морей
1. Биогеографическое районирование Мирового океана.
2. Ареалы морских животных и растений, реликты фауны.
3. Биологическая структура океана.
4. Концепции биологической структуры океана.
1.Биогеографическое районирование Мирового океана.При биогеографическом районировании Мирового океана, которое носит зональный характер, А. Г. Воронов (1987) использовал схему зоогеографического районирования, предложенную В. Г. Гептнером (1936). В результате были выделены семь биогеографических областей: Арктическая, Бореаль-но-Тихоокеанская, Бореально-Атлантическая, Тропико-Индо-Тихоокеанская, Тропико-Атлантическая, Нотально-Антарктическая (Субантарктическая) и Антарктическая (рис. 6).
Эти области являются общими для неритической и океанической водной среды, но не учитывают своеобразие жизни в бентали и пелагиали, других экологических зонах Мирового океана. В частности, в зоогеографическом районировании Мирового океана по донной фауне материковая отмель разделена на три царства: холодных и умеренных морей Северного полушария, тропическое, холодных и умеренных морей Южного полушария, а абиссаль на три области: Тихоокеанско-Северо-Индийскую, Атлантическую и Антарктическую.
1. Арктическая область. Ее южная граница между Северной Америкой и Европой проходит от острова Ньюфаундленд в направлении архипелага Шпицберген к северо-западной оконечности Кольского полуострова, а между Азией и Северной Америкой — по северной части Берингова моря. В области постоянно низкая температура воды (около 0°С), соленость ниже средней для океана (результат таяния льда и выноса пресных вод реками). Зимой и летом преобладающая часть акватории покрыта ледовым панцирем, встречаются полыньи и разводья. Растительность более холодных вод отличается преобладанием бурых водорослей, а теплых — зеленых. Красные водоросли представлены сравнительно равномерно. Летом у кромки тающих льдов образуется массовое скопление планктона, которое определяет богатое разнообразие водных обитателей от крылоногих моллюсков лимацин до нарвалов и белух (из китообразных), а также чаек и чистиков. Среди чаек эндемична белая, из чистиков характерен люрик. Гренландский кит, обильный раньше, почти полностью истреблен. Из тюленей обычны нерпа, морской заяц, хохлач и морж. Круглый год по льдам бродит белый медведь. Из рыб характерны сайка и навага. Из беспозвоночных много бокоплавов и равноногих, которые достигают здесь наибольшего разнообразия.
2, 3. Бореально-Тихоокеанская (Бореально-Пацифшеская) и Бореально-Атлантическая области. Эти области имеют значительное сходство. Им свойственны резкие сезонные колебания температуры воды – от 3 до 15° и более. Большинство организмов, обитающих здесь, эвритермны. По световому режимубореальные области океана отличаются от Арктической круглогодичной ежесуточной сменой дня и ночи, что позволяет растительным организмам вегетировать большую часть года. Границы областей неодинаковы для организмов, обитающих в пелагиали на разных глубинах. Если в районировании учитывать сублиторальную (от уровня воды до глубины, где есть растительность) фауну, то южная граница пройдет между 30 и 40° с. ш. и почти совпадет со среднегодовой изотермой 15°С. Система теплых и холодных течений у восточных берегов, как Северной Америки, так и Азии клинообразно суживает акваторию областей. По видовому разнообразию бореальные флора и фауна значительно превосходят арктическую, но уступают тропической. Наибольшее обилие организмов приурочено к литорали (затапливаемая прибрежная часть) и сублиторали.
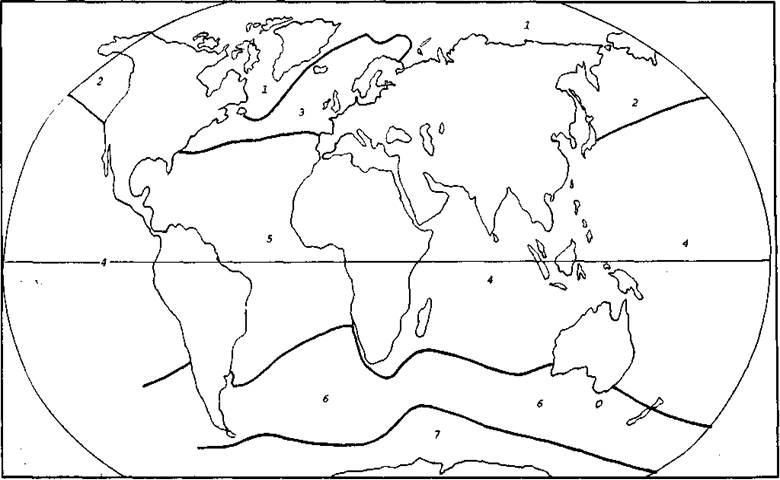
Рис. 6. Биогеографическое районирование Мирового океана (по Воронову, 1987):
Области: 1–Арктическая, 2 – Бореаль-но-Тихоокеанская, 3 – Бореально-Атлантическая, 4 – Тропико-Индо-Тихоокеанская, 5 – Тропико-Атлантическая, 6 – Нотально-Антарктическая (Субантарктическая), 7 – Антарктическая.
Для Бореально-Тихоокеанской области особенно характерны бурые водоросли(макроцистис и нереоцистис). Среди моллюсков обычны устрицы, мидии, морские гребешки, тихоокеанский кальмар и осьминог Дофлейна. Ракообразные представлены вислоногими (каланус тихоокеанский) и десятиногими раками (камчатский краб, креветки чилимы). Из иглокожих наиболее распространен трепанг. Рыбы (кета, горбуша, чавыча, нерка, иваси и др.) имеют большое промысловое значение. Из птицособенно много чистиковых. Разнообразны млекопитающие: морской котик, сивуч, калан, японский кит, серый кит, дельфин, белокрылая морская свинья.
В Бореально-Атлантической области обильны бурые водоросли (ламинария, алярия, фукусы) и красные(анфельция). Ракообразныепредставлены вислоногими раками, креветками, лангустами, омарами, рыбы – треской, пикшей, сайдой, кефалью, камбалой и зубаткой. Среди птиц много чаек, чистиков, кайр, гагарок, ластоногих– хохлачей, серых и гренландских тюленей, китообразных – гринд.
Некоторые представители фауны (серый дельфин и обыкновенная морская свинья, обыкновенный тюлень, чистики, сельдевая акула, морские сельди, треска и др.) имеют амфибореальное распространение у берегов Евразии и Северной Америки. Они отсутствуют в морях, омывающих северные побережья этих континентов. Л. С. Берг объяснил эту разорванность ареалов тем, что температура воды полярных морей в плиоцене была значительно более высокой и многие виды морских животных могли проникнуть из Северной Атлантики через полярные моря, омывающие северное побережье Евразии, в северную часть Тихого океана, и наоборот. Наступившее в антропогене похолодание вызвало резкое понижение температуры в полярных морях, и многие виды вымерли. В более южных широтах понижение температуры воды было не столь значительным, что способствовало сохранению формирующейся амфибореальной фауны.
4, 5. Тропика-Атлантическая и Тропика-Индо-Тихоокеанская области.Для них также характерны общие черты природных условий и биоты.
Прежде всего, постоянно высокая температура поверхностных слоев воды (выше 20 °С) с незначительными годовыми колебаниями (не более 2°), а также контрастные температурные различия между поверхностными и глубинными горизонтами. Северная граница этих областей совпадает с годовой изотермой воды 15 °С, а южная (в Южном полушарии) – 17°С. Только в этих областях распространены сообщества мангров и коралловых рифов. Из водорослей следует отметить саргассовые (саргассум и турбинария). В водах тропических областей обитает подавляющее большинство видов планктонныхфораминифер, крылоногих и киленогих моллюсков, сифонофор, кольчатых червей, сальп и аппендикулярий. В тропических областях обычны крупные акулы, летучие рыбы, меч-рыбы, парусники, скаты, морские черепахи, морские змеи, лангусты, жемчужницы. С тропическими морями связаны птицы фаэтоны и фрегаты, большую часть жизни проводящие в поисках пищи над водной поверхностью. Из млекопитающихобычны белобрюхие тюлени, кашалоты, дюгони и ламантины. Хотя в целом фауна тропических областей океана отличается большим разнообразием, она имеет значительно меньшую, чем в других поясах, численность отдельных видов. Только в районах апвеллингов наблюдаются массовые скопления животных.
Нотально-Антарктическая область. По условиям существования жизни эта область сходна с бореальными. Для нее характерны те жерезкие сезонные колебания температуры, подъемы глубинных вод, обогащенных биогенными веществами, и обилие жизни. Кроме биполярно распространенных представителей флоры и фауны (бурые водоросли, несколько видов китов, котики, обыкновенные тюлени, кильки, сардины и др.) следует отметить: из млекопитающих – гривистого сивуча, южного котика, южного и карликового китов, морского слона и морского леопарда, из рыб – представителей семейства нототениевых, а птиц – королевского альбатроса.
7. Антарктическая область. Как и Арктической, ей свойственны постоянно низкие температуры воды и развитие ледового покрова. Кроме того, условия существования жизни ограничены многочисленными айсбергами, которые, как бы перепахивая приливно-отливную полосу, уменьшают обилие жизни в литорали.
Антарктические воды более благоприятны для живых организмов, чем суша, и по этой причине их фауна несравненно разнообразнее. Морские беспозвоночные – криль (планктонные ракообразные) летом в поверхностных слоях воды образуют огромные скопления, служащие кормом для ряда видов рыб, птиц и млекопитающих. Из рыб распространено семейство белокровных щук. Летом многочисленны буревестники и поморники, нередко встречаются крачки, альбатросы и качурки. Наиболее типичные представители области — пингвины. У берегов Антарктиды, близ островов и среди дрейфующих льдов обитают настоящие тюлени (Уэдделла, Росса, крабоед, морской леопард, морской слон). Довольно многочислен морской котик. Массовое скопление криля привлекает большие стада китов (синего, финвала, горбача, сейвала, полосатика и др.). Встречаются кашалоты, касатки и бутылконосы. Своеобразна донная фауна Антарктической области. Обильны губки и иглокожие. Масса медуз достигает 156 кг.
К сожалению, флора и фауна Мирового океана в значительной мере испытали на себе разрушительное воздействие антропогенного фактора. Не только уменьшилась численность их представителей, но и полностью уничтожены отдельные виды, загрязнены нефтью, нефтепродуктами, бытовыми стоками и разнообразными токсичными веществами промышленного происхождения воды.
Дата добавления: 2015-07-18 ; просмотров: 3914 ; ЗАКАЗАТЬ НАПИСАНИЕ РАБОТЫ
Источник
384 Биогеографическое районирование океана
При биогеографическом районировании океана, основанном на изучении распространения и распределения организмов, проводится разделение океана на регионы (акватории) различного ранга: области, подобласти, провинции. Районирование проводят или по таксономическому разнообразию биот, или по разнообразию структуры образуемых ими сообществ. При установлении границ используются индикаторные виды.
А. Ортман (1896) впервые произвел районирование для литоральной (литораль + сублитораль + часть батиали), абиссальной и пелагической фаун отдельно, так как проведение единых зоогеографических границ для всей толщи океана невозможно. Условия расселения организмов в абиссали, в сублиторали и в пелагиали совсем иные, и поэтому не могут быть общими и зоогеографические границы. Эти подходы не утратили своей значимости и поныне.
Литораль весьма насыщена жизнью. В настоящее время здесь выделяются две области: Бореальная и Тропическая. Одной из ярких особенностей Тропической области является распространение коралловых рифов и богатейшей сопутствующей им фауны, в том числе множества своеобразных коралловых рыб, моллюсков, иглокожих, актиний. Насколько богата эта фауна, можно судить по
 Рис. 88. Рыбопродуктивность Мирового океана в кг/км 2(на 1985 г.) (П.А.Моисеев, 1989): 1 — > 3000; 2 — > 1000; 3-> 500; 4-> 100; 5 — > 50; 6 — > 10; 7 — 2 .
Рис. 88. Рыбопродуктивность Мирового океана в кг/км 2(на 1985 г.) (П.А.Моисеев, 1989): 1 — > 3000; 2 — > 1000; 3-> 500; 4-> 100; 5 — > 50; 6 — > 10; 7 — 2 .
Известно уже несколько десятков подобных излияний в Тихом (там их больше всего), Атлантическом, Индийском и Северном Ледовитом океанах. Число уже описанных эндемичных видов приближается к 250. Среди них представлены и эндемичные таксоны
 Рис. 90. Зоны разломов (1) и районы обнаружения сообществ хемобиоса (2) в Мировом океане (А.М.Сагалевич, Л.И.Москалев, 1991)
Рис. 90. Зоны разломов (1) и районы обнаружения сообществ хемобиоса (2) в Мировом океане (А.М.Сагалевич, Л.И.Москалев, 1991)
высокого ранга, вплоть до класса. Около трети видов приходится на долю многощетинковых червей (полихет), особенно из живущих в трубках «помпейских червей». Столь же разнообразны брюхоногие моллюски. Далее в порядке убывания числа видов следуют веслоногие рачки, десятиногие раки, особый класс — вестиментиферы, двустворчатые моллюски и др. (рис. 91).
Исследования экосистем гидротерм обогатили науку открытием вестиментифер: своеобразных, похожих на червей, сидящих в трубках животных, относящихся к новому самостоятельному классу. Вестиментиферы не имеют кишечника и существуют за счет симбиотических бактерий, потребляющих соединения серы.
Гидротермальные морские экосистемы существуют за счет процессов хемосинтетического образования органических веществ. Напомним, что более 100 лет назад выдающийся русский микробиолог С. Н. Виноградский (1856-1953) открыл хемосинтез — образование некоторыми бактериями органических веществ из диоксида углерода не за счет солнечной энергии, как при фотосинтезе, а за счет энергии, получаемой при окислении восстановленных неорганических соединений. За последние 35 лет активное исследование дна Мирового океана позволило выявить в многочисленных зонах спрединга и субдукции литосферных плит, теплового разложения газогидратов, выхода грунтовых вод и т. п. на самых разных уровнях — от уреза воды до глубин свыше 6 км — множество
 Рис. 91. Типичное сообщество макрофауны гидротермального вента (моллюски, крабы, вестиментиферы и др. вокруг источников серосодержащих подземных вод на дне океана) (Г. Макдональд и др., 1985)
Рис. 91. Типичное сообщество макрофауны гидротермального вента (моллюски, крабы, вестиментиферы и др. вокруг источников серосодержащих подземных вод на дне океана) (Г. Макдональд и др., 1985)
участков, населенных специфическими сообществами донных организмов, существующих на основе хемосинтеза. Таким образом, все население Мирового океана может быть разделено по происхождению потребляемого им органического вещества на фотобиос и хемобиос. Хемобиос в пищевом отношении не зависит от энергии Солнца, но зависит от нее как от источника кислорода: поступающий в океанские глубины кислород имеет фотосинтетическое происхождение.
Сообщества хемобентоса на дне океана всегда крайне ограничены по площади — в масштабе океана это точки. Обнаружить их обычными гидробиологическими методами — с помощью тралов и дночерпателей — никому не удалось за 100 лет интенсивных биологических исследований океана.
Что же касается биогеографии «гидротермального вента» — так называют экологи хемотрофные донные сообщества, — то здесь сделаны только первые шаги, выявившие привязанность таких экосистем к разломам на дне океана и к зонам выхода на поверхность богатых серой низкотемпературных вод.
Гидротермальные источники в море — явление не столь уж редкое; есть они, например, и в районе Курильских островов, в том числе и в сравнительно мелководных заливах. В 1985 г. экспедиция на гидрографическом судне «Таймыр» обнаружила такое своеобразное сообщество организмов в почти изолированной от океана бухте Кратерной на о. Янкича (Средние Курилы). Бухта Кратерная представляет собой затопленный кратер потухшего вулкана, который соединяется с океаном очень мелким проливом. Из гидротермальных источников, расположенных на берегу и под водой, в бухту постоянно поступают горячие и сильно минерализованные воды, несущие серу и ее соединения; со дна бухты пузырями поднимается сероводород. «Плотность населения» на дне бухты составляет сотни и тысячи особей, а биомасса — несколько килограммов на 1 м 2 . По мнению А. В.Жирмунского, основу существования этой уникальной экосистемы составляют, по-видимому, сбалансированные процессы интенсивного бактериального хемосинтеза и фотосинтеза водорослей. Еще одна отличительная ее особенность — высокая скорость и отлаженность круговорота биогенных веществ, благодаря которой поддерживается стабильность системы.
Источник