- Первичная продуктивность Мирового океана
- Первичная продуктивность мирового океана
- Изменчивость и моделирование первичной продукции в Карском море
- Изменчивость первичной продукции в Черном море
- Использование флуоресцетного метода в биоокеанологии
- Методы определения и оценки первичной продукции
- Оценка первичной продукции в морских экосистемах
- Флуоресцентный метод изучения параметров первичной продуктивности
- Modelling Kara Sea phytoplankton primary production: Development and skill assessment of regional algorithms
- Моделирование первичной продукции Карского моря.
- Он-лайн представление параметров, определяющих первичную продуктивность Карского моря.
- Сезонная изменчивость первичной продукции фитопланктона Карского моря по спутниковым данным.
- Первичная морская продукция — Marine primary production
- СОДЕРЖАНИЕ
- Морские первичные производители
- Цианобактерии
- Биологические пигменты
- Хлоропласты
- Микробный родопсин
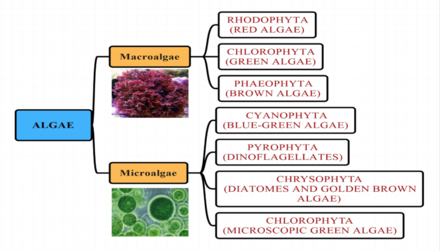
- Морские водоросли
- Группы водорослей
- Зеленые водоросли
- Красные водоросли
- Бурые водоросли
- Диатомеи
- Кокколитофориды
- Динофлагеллята
- Миксотрофные водоросли
- Другие группы
- По размеру
- Микроводоросли
- Макроводоросли
- Морские растения
- Мангровые заросли
- Морские травы
- Стехиометрия
Первичная продуктивность Мирового океана
Первичная продукция – скорость образования органического вещества в процессе фотосинтеза. Основная часть первичной продукции Мирового океана создается фитопланктоном. Существуют два аспекта изучения первичной продукции океанического фитопланктона – биогеохимический и экологический.
Биогеохимический аспект связан с выяснением причин изменчивости содержания углекислого газа в океане и атмосфере. Океан – важнейшее звено в глобальном балансе СО2. Общее количество СО2 в океане в 50 раз больше, чем в атмосфере. Даже небольшие изменения обменного потока углекислого газа между океаном и атмосферой могут существенно изменить концентрацию в ней СО2.
Экологический аспект связан с изучением закономерностей формирования первичной продукции и ее роли в функционировании морских экосистем. Оценка величины первичной продукции морей, океанических акваторий и Мирового океана в целом предполагает исследование закономерностей ее формирования, пространственно-временной изменчивости, физиологического состояния фитопланктонного сообщества, создание продукционных моделей, использование спутниковых данных, исследования переноса новообразованного в процессе фотосинтеза органического вещества по пищевым цепям от низших трофических уровней к его конечным потребителям.
Величина первичной продукции служит показателем биопродуктивности морских экосистем в целом.
Решением этих вопросов в Институте океанологии им П. П. Ширшова РАН занимается межлабораторная группа, имеющая большой опыт экспедиционных работ в различных районах Мирового океана.
Источник
Первичная продуктивность мирового океана
Демидов А.Б.
Первичная продукция океанического фитопланктона: изменчивость и глобальные оценки.
Соавторы: Мошаров С.А., Гагарин В.И.
Изменчивость и моделирование первичной продукции в Карском море
Демидов А. Б., Мошаров С. А.
Первичная продукция Карского моря: изменчивость и моделирование
Изменчивость первичной продукции в Черном море
А.Б.Демидов, В.И.Ведерников
Оценки первичной продукции Черного моря и ее сезонная изменчивость.
Использование флуоресцетного метода в биоокеанологии
С.А.Мошаров
Использование флуоресцентного метода в биоокеанологии.
Методы определения и оценки первичной продукции
С.А.Мошаров, А.Б.Демидов
Современные методы определения и оценки первичной продукции в морях и океанах.
Оценка первичной продукции в морских экосистемах
А.Б.Демидов, С.А.Мошаров
Оценка первичной продукции в морских экосистемах
Флуоресцентный метод изучения параметров первичной продуктивности
С.А.Мошаров
Флуоресцентный метод изучения распределения хлорофилла и процессов первичной продукции в водной среде.
Modelling Kara Sea phytoplankton primary production: Development and skill assessment of regional algorithms
Andrey B. Demidov, Oleg V. Kopelevich, Sergey A. Mosharov,
Sergey V. Sheberstov, Svetlana V. Vazyulya
Modelling Kara Sea phytoplankton primary production: Development and skill assessment of regional algorithms
Моделирование первичной продукции Карского моря.
Демидов А.Б.
Моделирование первичной продукции Карского моря.
Он-лайн представление параметров, определяющих первичную продуктивность Карского моря.
Мошаров С.А., Демидов А.Б., Метальников А.А., Шеберстов С.В.
Он-лайн представление параметров, определяющих первичную продуктивность Карского моря.
Сезонная изменчивость первичной продукции фитопланктона Карского моря по спутниковым данным.
А.Б.Демидов, С.В.Шеберстов, В.И.Гагарин, П.В.Хлебопашев
Сезонная изменчивость первичной продукции фитопланктона Карского моря по спутниковым данным.
Источник
Первичная морская продукция — Marine primary production

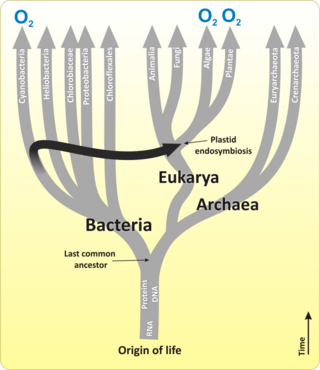
Первичное производство в море — это химический синтез в океане органических соединений из атмосферного или растворенного углекислого газа . В основном это происходит в процессе фотосинтеза , в котором в качестве источника энергии используется свет, но также в процессе хемосинтеза , который использует окисление или восстановление неорганических химических соединений в качестве источника энергии. Почти вся жизнь на Земле прямо или косвенно зависит от первичного производства . Организмы, ответственные за первичную продукцию, называются первичными продуцентами или автотрофами .
Большая часть первичной морской продукции производится разнообразным набором морских микроорганизмов, называемых водорослями и цианобактериями . Вместе они образуют основных первичных производителей в основе пищевой цепи океана и производят половину мирового кислорода. Морские первичные продуценты поддерживают почти все виды морских животных, производя почти весь кислород и пищу, которые необходимы морским животным. Некоторые морские первичные производители также являются инженерами экосистем, которые изменяют окружающую среду и обеспечивают среду обитания для других морских обитателей.
Первичное производство в океане можно сравнить с первичным производством на суше. В глобальном масштабе и океан, и суша производят примерно одинаковое количество первичной продукции, но в океане первичная продукция происходит в основном за счет цианобактерий и водорослей, а на суше — за счет сосудистых растений .
Морские водоросли включают в себя в основном невидимые и часто одноклеточные микроводоросли , которые вместе с цианобактериями образуют фитопланктон океана , а также более крупные, более заметные и сложные многоклеточные макроводоросли, обычно называемые водорослями . Водоросли встречаются в прибрежных районах, обитают на дне континентальных шельфов и вымываются в приливных зонах . Некоторые водоросли плывут вместе с планктоном в освещенных солнцем поверхностных водах ( эпипелагиали ) открытого океана.
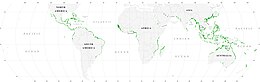
Еще в силурийский период часть фитопланктона превратилась в красные , коричневые и зеленые водоросли . Затем эти водоросли вторглись на сушу и начали превращаться в наземные растения, которые мы знаем сегодня. Позже в меловом периоде некоторые из этих наземных растений вернулись в море в виде мангровых зарослей и морских трав . Они встречаются вдоль побережья в приливных районах и в солоноватой воде эстуариев . Кроме того, некоторые виды морских водорослей, например водоросли, можно найти на глубине до 50 метров как на мягком, так и на твердом дне континентального шельфа.
| Часть серии обзоров по | ||||||
| морская жизнь | ||||||
|---|---|---|---|---|---|---|
 | ||||||
| ||||||
 Портал морской жизни Портал морской жизни | ||||||
| Часть серии о | |||
| Углеродный цикл | |||
|---|---|---|---|
 | |||






























| Внешнее видео | |
|---|---|
 Как возникла многоклеточность? — Путешествие в микрокосмос Как возникла многоклеточность? — Путешествие в микрокосмос |
Стехиометрия
Стехиометрии (измерение химических реагентов и продуктов ) первичной продукции в поверхности океана играет решающую роль в круговороте элементов в мировом океане. Соотношение между элементами углерода (С), азот (N) и фосфора (Р) в экспортируемой органического вещества выражены в терминах C: N: Р помогает определить , сколько атмосферного углерода является поглощенных в глубоком океане по отношению к наличие ограничивающих питательных веществ . В геологическом масштабе времени соотношение N: P отражает относительную доступность нитрата по отношению к фосфату , оба из которых поступают извне из атмосферы посредством фиксации азота и / или с континентов через реку и теряются в результате денитрификации и захоронения. В более коротких временных масштабах средняя стехиометрия экспортируемого органического вещества в виде твердых частиц отражает элементарную стехиометрию фитопланктона с дополнительным влиянием биологического разнообразия и вторичной переработки органического вещества зоопланктоном и гетеротрофными бактериями. Перед лицом глобальных изменений понимание и количественная оценка механизмов, которые приводят к изменчивости соотношений C: N: P, имеют решающее значение для точного прогноза будущего изменения климата.
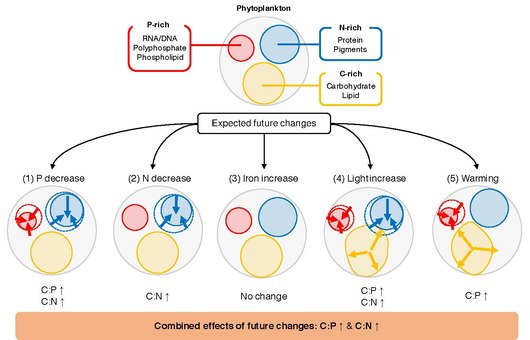
Ключевой нерешенный вопрос заключается в том, что определяет C: N: P отдельного фитопланктона. Фитопланктон растет в верхнем освещенном слое океана , где количество неорганических питательных веществ, света и температуры варьируется в пространстве и во времени. Лабораторные исследования показывают, что эти колебания вызывают реакции на клеточном уровне, посредством чего клетки изменяют распределение ресурсов, чтобы оптимально адаптироваться к окружающей среде. Например, фитопланктон может изменять распределение ресурсов между богатым P биосинтетическим аппаратом, богатым азотом светособирающим аппаратом и запасами накопления энергии, богатыми углеродом. При типичном сценарии будущего потепления ожидается, что глобальный океан будет претерпевать изменения в доступности питательных веществ, температуре и освещенности . Эти изменения, вероятно, окажут глубокое влияние на физиологию фитопланктона, и наблюдения показывают, что конкурентоспособные виды фитопланктона могут акклиматизироваться и адаптироваться к изменениям температуры, освещенности и питательных веществ в десятилетних временных масштабах. Были проведены многочисленные лабораторные и полевые эксперименты, изучающие взаимосвязь между соотношением C: N: P фитопланктона и факторов окружающей среды. Однако сложно синтезировать эти исследования и обобщить реакцию фитопланктона C: N: P на изменения факторов окружающей среды. В отдельных исследованиях используются различные наборы статистических анализов для характеристики влияния факторов окружающей среды на элементные отношения, начиная от простого t-теста и заканчивая более сложными смешанными моделями, что затрудняет сравнение результатов между исследованиями. Кроме того, поскольку изменения признаков, вызванные окружающей средой, обусловлены сочетанием пластичности ( акклиматизации ), адаптации и жизненного цикла, стехиометрические реакции фитопланктона могут быть разными даже среди близкородственных видов.
Мета-анализ / систематический обзор — это мощная статистическая основа для синтеза и интеграции результатов исследований, полученных в результате независимых исследований, и для выявления общих тенденций. Основополагающий синтез Гейдера и Ла Роша в 2002 г., а также более поздняя работа Перссона и др. в 2010 году показал, что C: P и N: P могут варьироваться в 20 раз между клетками с высоким и ограниченным содержанием питательных веществ. Эти исследования также показали, что соотношение C: N может быть умеренно пластичным из-за ограничения питательных веществ. В метааналитическом исследовании Hillebrand et al. в 2013 году подчеркнули важность скорости роста в определении элементной стехиометрии и показали, что отношения C: P и N: P снижаются с увеличением скорости роста. В 2015 году Ивон-Дюроше и др. исследовал роль температуры в модулировании C: N: P. Хотя их набор данных ограничивался исследованиями, проведенными до 1996 года, они показали статистически значимую взаимосвязь между C: P и повышением температуры. MacIntyre et al. (2002) и Thrane et al. (2016) показали, что освещенность играет важную роль в управлении оптимальным соотношением C: N и N: P в клетках. Совсем недавно Морено и Мартини (2018) представили исчерпывающий обзор того, как условия окружающей среды регулируют клеточную стехиометрию с физиологической точки зрения.
Элементная стехиометрия морского фитопланктона играет решающую роль в глобальных биогеохимических циклах благодаря своему влиянию на круговорот питательных веществ, вторичную продукцию и экспорт углерода. Хотя на протяжении многих лет проводились обширные лабораторные эксперименты для оценки влияния различных факторов окружающей среды на элементный состав фитопланктона, всеобъемлющая количественная оценка процессов все еще отсутствует. Здесь ответы соотношений P: C и N: C морского фитопланктона были синтезированы на пять основных факторов (неорганический фосфор, неорганический азот, неорганическое железо, освещенность и температура) с помощью метаанализа экспериментальных данных по 366 экспериментам из 104 журнальные статьи. Эти результаты показывают, что реакция этих соотношений на изменения в макронутриентах одинакова во всех исследованиях, где увеличение доступности питательных веществ положительно связано с изменениями соотношений P: C и N: C. Результаты показывают, что фитопланктон эукариот более чувствителен к изменениям макроэлементов по сравнению с прокариотами, возможно, из-за их большего размера клеток и их способности быстро регулировать паттерны экспрессии генов. Эффект освещенности был значительным и постоянным во всех исследованиях, где увеличение освещенности уменьшало как P: C, так и N: C. Отношение P: C значительно снижалось при нагревании, но реакция на изменения температуры была неоднозначной в зависимости от режима роста культуры и фазы роста во время сбора урожая. Наряду с другими океанографическими условиями субтропических круговоротов (например, низкая доступность макроэлементов) повышенная температура может объяснить, почему P: C постоянно низкий в субтропических океанах. Добавление железа не изменяло систематически ни P: C, ни N: C.
Источник